Биометрический метод определения пола осетровых, в частности - Русского осетра Acipenser gueldenstaedti (Acipenseridae) Азовской популяции
На примере русского осетра Acipenser gueldenstaedti азовской популяции показана возможность биометрического определения пола осетровых рыб. Предложена схема промеров головы, позволяющая Эффективно проводить прижизненную оценку пола. Получено дискриминантное уравнение, позволяющее в достаточно широком размерно-возрастном диапазоне и вне зависимости от стадии зрелости гонад идентифицировать пол особей русского осетра.
Список методов определения половой принадлежности у осетровых рыб (Acipenseridae) включает: анатомические (биопсийные и эндоскопический), физиолого-биохимические, эндокринологические и ультразвуковой диагностики (УЗИ). Этот перечень будет, вероятно, расширяться, что обусловлено не только актуальностью определения пола у осетровых, но и наличием серьезных недостатков у вышеуказанных методов. К числу наиболее распространенных и представляющих наибольший интерес методов относятся биопсия гонад и УЗИ.
Неоспоримым преимуществом биопсийного метода, наиболее широко применяемого в рыбоводстве при оценке половой принадлежности особей, является возможность получения дополнительной информации о зрелости гонад, вплоть до оценки степени поляризации ооцитов. Однако извлечение специальным щупом фрагмента гонады на I и II стадиях зрелости - процедура довольно трудная и не всегда результативная. Кроме того, в гонадах рыб жировой ткани бывает значительно больше, чем генеративной, что еще больше затрудняет взятие нужной пробы. Вследствие этих обстоятельств данный способ практически применим не ранее достижения гонадами П-Ш и Ш стадий зрелости. К недостаткам метода следует отнести и его травматичность.
Появление современного метода ультразвуковой диагностики, позволяющего быстро (в чем его главное, на наш взгляд, преимущество перед другими методами) и безболезненно сканировать внутренние органы, привлекло внимание многих специалистов и обеспечило перспективы использования УЗИ в ветеринарии и рыбоводстве (Karlsen, Holm, 1994). Однако широкому распространению этого метода препятствует высокая стоимость работ и, прежде всего, оборудования. Следует также отметить, что эхографическое изображение структуры гонад отнюдь не адекватно визуальному, поэтому качественная диагностика пола требует серьезной подготовки оператора. При этом результативность определения пола (правильность интерпретации эхографическо-го изображения), как и при использовании биопсин ного метода, полностью зависит от стадии зрелосп гонад и даже при наличии тренированного персона ла остается не очень высокой, в среднем 80% (Williot. 2002) и 80-90% (Billard, 2002).
Вполне конкурентоспособным мог бы стать биометрический метод идентификации пола по количественным признакам, но предлагаемые для осетровых схемы промеров пластических и меристических признаков (Соколов, Кашин. 1965; Крылова, Соколов, 1981), в основе которых лежит традиционная для отечественных ихтиологов схема Правдина (1966), оказались явно непригодными для оценки полового диморфизма. Даже применение аналитических методов с высокой разрешающей способностью (например, дискри-минантного анализа) оказалось неэффективным, хотя при этом была показана достаточно высокая (с ошибкой в классификации 1.6%) возможность различать отдельных особей азовской и каспийской популяций севрюги Acipenser stellatus (Цвет-ненко, 1993). Традиционная схема промеров оказалась малоэффективной и в других случаях. Так. идентификация пола (с использованием дискри-минантного анализа) у белого толстолобика Ну-pophthalmichthys molithx (Волчков и др., 1990) была возможна, но с довольно большой ошибкой (13.3%). Попытка оценки половой принадлежности осетровых по форме урогенитальных отверстий (Vecsei et al., 2003) также показала низкую результативность (ошибка классификации 29%).
Проявления полового диморфизма были отмечены при краниологических исследованиях костистых рыб (Васильева, 1997). Оригинальная схема краниометрических промеров (отделов и отдельных костей дермокраниума), разработанная Васильевой на основе музейных коллекций осетров, была также использована для морфологической оценки андрогенетических гибридов русского A. gueldenstaedti и персидского A. persicus осетров (Васильева и др., 2001).
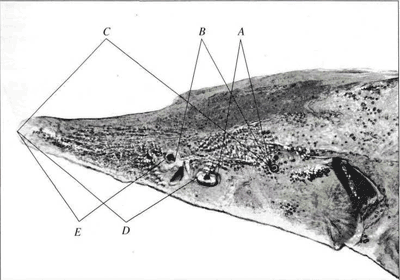
Рис. 1.Схема промеров - латеральная проекция.
Вышеизложенные и полученные нами предварительные результаты определили необходимость в большей детализации схемы промеров головы осетровых как более перспективной для определения пола и таксономической идентификации. После предварительной оценки, включающей традиционные промеры головы (отдельные из них вошли в предложенную схему), были отобраны пластические признаки, отвечающие следующим требованиям: 1 - достаточно изменчивые для достоверного определения как половой, так и таксономической идентификации отдельных особей; 2 - универсальные, т.е. поддающиеся оценке у всех представителей семейства осетровых; 3 — поддающиеся измерению в широком диапазоне размеров особей; 4 - точно и быстро измеряемые.
МАТЕРИАЛ И МЕТОДИКА
Материал для исследования получен в 2004-2005 гг. на экспериментальном участке Южного филиала Федерального селекционно-генетического центра рыбоводства и в Южном производственном осетрово-рыбоводном центре (Республика Адыгея). Для биометрических измерений использовали случайную выборку из 36 самок и 36 самцов русского осетра A. gueldenstaedti, какполученных (генерации 1995-2002 гг.) и выращенных в условиях искусственного содержания (возраст от 2+ до 10+), так и выловленных в 2004-2005 гг. в Азовском море.
Длина исследуемых рыб от конца рыла до конца средних лучей хвостового плавника, промер АС по традиционной схеме (Правдин, 1966), варьировала в пределах 54.0-105.0 (самцы) и 61.0-160.0 см (самки). Ее измеряли с точностью до 1 мм, используя стандартную миллиметровую ленту. Пол особей определяли при получении от них зрелых половых продуктов или (при I—III стадии зрелости гонад) оперативным методом (вскрытие рыб и определение пола по внешнему виду гонад).
Используемая нами схема (см. рис. 1-3: показана в виде примера на особи ленского осетра A. baerii) включала 17 промеров головы и прилегающих к ней частей тела между: брызгальцем и глазом (А), брызгальцем и передним носовым отверстием (В), брызгальцем и концом рыла (С), глазом и концом рыла (D), передним носовым отверстием и концом рыла (E), верхними краями жаберных крышек (F), брызгальцами (G), глазами (H), передними носовыми отверстиями (/), ключичными костями (У, М, N, О, Р), серединой соединения ключиц и концом рыла (К), серединой хрящевого свода рта и концом рыла (L), грудными плавниками (Q). Измерения проводили чертежным измерителем с точностью до 1 мм. Следует отметить, что в популяционных исследованиях обычно используют не только непосредственно измеряемые признаки, но и их комбинации, в частности, отношения (индексы), отражающие пропорции исследуемых объектов.
Рис. 2. Схема промеров - дорзальная проекция.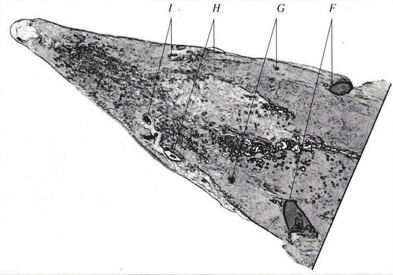
Рис. 3. Схема промеров - вентральная проекция.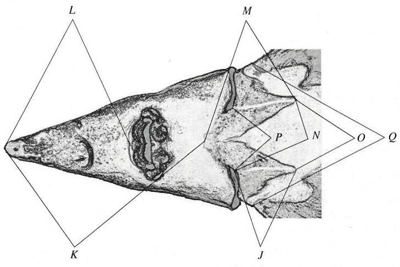
Было показано, что обе системы признаков имеют свои недостатки и преимущества (Андреев, Решетников, 1977), а замена исходных признаков индексами далеко не всегда оказывается полезной (Животовский, 1984). В ихтиологических работах используют систему индексов, прежде всего, для уменьшения влияния размеров особей. При этом величину промера относят к общей длине тела или к длине головы (для промеров головы), а не в виде всех возможных отношений промеров друг к другу. Очевидно, что при таком подходе наиболее ценные комбинации отношений, обладающие меньшей вариабельностью и лучшими дифференцирующими свойствами, могут оказаться просто не включенными в анализ данных. В нашей работе были использованы обе системы признаков совместно: система непосредственно измеряемых значений (17 промеров) и система индексов - в виде 136 всех возможных сочетаний отношений промеров.
Таблица 1. Статистическая оценка дискриминантной функции.
|
Коэффициент канонической корреляции |
χ2 |
U (Wilks lambda) |
F - аппроксимация |
dƒ - число степеней свободы |
Уровень значимости |
|
0.78 |
30.7 |
0.38 |
12.48 |
4 |
< 0001 |
Таблица 2. Результаты классификации половой принадлежности особей русского осетра Acipenser gueldenstaedti.
|
Пол |
Число рыб в группе, экз. |
Прогноз пола, экз. (%) |
Общее число неправильно классифицированных особей, % | |
|
самки |
самцы | |||
|
Обучающая подвыборка | ||||
|
Самки |
18 |
16 |
2 |
5.56 |
|
Самцы |
18 |
0 |
18 | |
|
Текстовая подвыборка | ||||
|
Самки |
18 |
16 |
2 |
8.33 |
|
Самцы |
18 |
17 |
1 | |
Следует отметить, что при проведении серии последовательных вычислений может произойти накопление погрешностей, поэтому в подобных случаях рекомендуется (Снедекор, 1961) брать на 2-3 знака больше необходимого в окончательном виде (в нашем случае - оценки параметров дискриминантной функции). Поэтому индексы рассчитывали с точностью до девятого знака.
При создании прогнозирующего уравнения применяли пошаговый дискриминантный анализ. При этом половину наблюдений исходной выборки (18 самцов и 18 самок) использовали в виде "обучающей" подвыборки, а остальные (18 самцов и 18 самок) - в виде "тестовой".
Для предварительного выбора "наилучшего" подмножества переменных применялись рекомендации Рао (1968) и Афифи, Эйзена (1982), а поскольку для двух групп дискриминантный анализ может рассматриваться и как процедура множественной регрессии, были также использованы предложения Мостеллера и Тьюки (1982) и Дрейпера и Смита (1987). При окончательном отборе переменных использовали обычно рекомендуемые (Дрей-пер, Смит, 1987: 24) величины для F включения (удаления): F0.95=4. Адекватность подобранной модели оценивали по графическому распределению остатков и статистики Дарбина-Уотcона (Афифи, Эйзен, 1982: 190; Дрейпер, Смит, 1987:246). Для вышеуказанных операций использовали пакет статических программ STATGRAPHICS Plus Version 5.1.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
В результате анализа были отобраны 4 "наилучших" переменных (/, Q, I/Q, Е/Н) и определены их коэффициенты, максимизирующие расстояние Махаланобиса - Δ2 (в данном случае его выборочное расстояние - D2) между полами. Остальные переменные не вносили значимого вклада в дискриминацию полов. Для проверки гипотезы Hо (об отсутствии различий между самками и самцами) использовались величины χ2 и F- аппроксимацию U - статистики. В обоих случаях гипотезу Hо отвергали на уровне значимости р < 0001 (табл. 1).
Критерием качества дискриминации является доля неверно классифицированных особей (или процент отнесения в "свою" группу). Известно, что выборочная оценка D2 расстояния Махаланобиса является смещенной. Поэтому нами был применен один из рекомендуемых методов (Афифи, Эйзен, 1982: 331), дающий несмещенные оценки, где наблюдения из первой подвыборки (обучающей) используются для создания дискриминантной функции, а особи второй подвыборки (тестовой) классифицируются для оценки точности прогноза согласно процедуре, полученной по первой подвыборке.
В нашем случае (табл. 2, рис. 4) ошибка классификации составила для обучающей подвыборки 5.56% (2 самки), а для тестовой 8.33% (2 самки и 1 самец). Так как различие в качестве результатов между подвыборками оказалось невелико, а в обоих случаях было правильно классифицировано более 90.0% особей, можно говорить, что используемая нами модель вполне удовлетворительно выдержала проверку и может быть использована для идентификации половой принадлежности особей.
В общем виде дискриминантное уравнение обычно представляется как:
D2 = a + b1X1+b2X2 + ... + bpXp,
где D2 - оценка выборочного расстояния Махаланобиса, а - константа, b1...bp - нестандартизиро-ванные коэффициенты, Х1.. .Хр — переменные, р -число переменных. Соответственно, полученное для определения пола уравнение можно представить в виде:
Рис. 4. График дискриминантной функции распределения самок и самцов русского осетра Acipenser gueldenstaedti.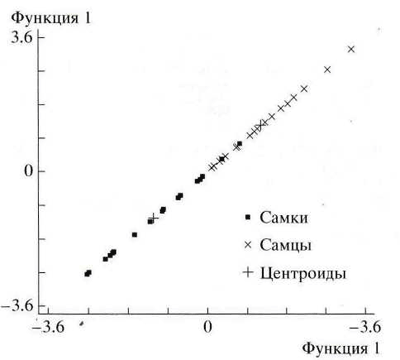
D2 = -36.7303 -0.696098I + 0.193362Q+ 101.344I/G+ 12.5249E/Q.
Оценивая вышеизложенные результаты в целом, следует отметить, что само проведение качественного дискриминантного анализа, конечно, требует определенных профессиональных навыков и знаний, но использование его результатов может быть доступно широкому кругу пользователей - ихтиологам, селекционерам и рыбоводам.
При наличии оценок параметров дискриминантной функции (нестандартизированных коэффициентов и константы) для определения знака функции (отрицательный — самки, положительный — самцы) необходим только программируемый калькулятор или карманный персональный компьютер. Время, необходимое для распознавания самок и самцов, ограничено только скоростью измерений (при определенных навыках - 30-40 с).
ЗАКЛЮЧЕНИЕ
Предложенная схема промеров с использованием дискриминантного анализа оказалась вполне эффективной для оценки полового диморфизма, а полученное дискриминантное уравнение позволяет достаточно быстро в широком размерно-возрастном диапазоне и вне зависимости от стадии зрелости гонад определять пол особей русского осетра.
Биометрический метод, конечно же, не лишен недостатков. Прежде всего, вследствие наличия краниологических различий между видами и подвидами у осетровых возникает необходимость иметь и отдельные уравнения для определения пола. Однако простота и эффективность метода являются безусловным преимуществом.
БЛАГОДАРНОСТИ
Выражаем сердечную признательность сотрудникам Южного филиала Федерального селекционно-генетического центра рыбоводства М.С. Чебанову и Ю.Н. Чмырь за поддержку и оказанную в работе помощь.
СПИСОК ЛИТЕРАТУРЫ
Андреев В.Л., Решетников С.Ю. 1977. Исследование внутривидовой морфологической изменчивости сига Corego-nus lavaretus (L.) методами многомерного статистического анализа // Вопр. ихтиологии. Т. 17. Вып. 5. С. 862-878.
Афифи А., Эйзен С. 1982. Статистический анализ: подход с использованием ЭВМ. М.: Мир, 488 с.
Васильева Е.Д. 1997. Размерная изменчивость и половой диморфизм костистых рыб: определяющие факторы и основные тенденции // Вопр. ихтиологии. Т. 37. № 3. С. 293-306.
Васильева Е.Д., Грунина А.С Рекубратский А.В. 2001. Характер проявления некоторых морфологических признаков у андрогенетических ядерно-цитоплаз-матических гибридов персидского Acipenser persicus и русского A. gueldenstaedti осетров в постларвальном онтогенезе // Вопр. ихтиологии. Т. 41. № 4. С. 530-537.
Волчков Ю.А., Решетников С.И., Илясова В.А. и др. 1990. Методические указания по оценке темпа полового созревания растительноядных рыб. М.: ВНИИПРХ, 33 с.
Дрейпер Н., Смит Г. 1987. Прикладной регрессионный анализ. Кн. 2. М.: Финансы и статистика, 351 с.
Животовский Л.А. 1984. Интеграция полигенных систем в популяциях (проблемы анализа комплекса признаков). М.: Наука, 184 с.
Крылова В.Д., Соколов Л.И. 1981. Морфологические исследования осетровых рыб и их гибридов (методические рекомендации). М.: ВНИРО, 49 с.
Мостеллер Ф., Тьюки Дж. 1982. Анализ данных и регрессия. Вып. 2. М.: Финансы и статистика, 239 с.
Правдин И.Ф. 1966. Руководство по изучению рыб. М.: Пищ. пром-сть, 376 с.
Рао С.Р. 1968. Линейные статистические методы и их применения. М.: Наука, 548 с.
Снедекор Дж.У. 1961. Статистические методы в применении к исследованиям в сельском хозяйстве и биологии. М.: Сельхозгиз, 502 с.
Соколов Л.И.. Кашин С.М. 1965. Сравнительный анализ некоторых морфологических показателей у популяций рыб сибирского осетра различных водоемов // Вестн. Моск. ун-та. № 3. С. 13-18.
Цветненко Ю.Б. 1993. Эффективность и генетические последствия интродукции севрюги Acipenser stellatus из Каспийского в Азовский бассейн // Вопр. ихтиологии. Т. 33. № 3. С. 382-387.
BillardR. 2002. Genetique // Esturgeons et caviar. Paris: Tec et Doc. (Aquaculture-Pisciculture). P. 91-106.
Karlsen 0., Holm ./. 1994. Ultrasonograph, a non-invasive method for sex determination in cod (Gadus morhua) // J. Fish Biol. V. 44. P. 965-971.
Vecsei P., Litvak M.K., Noakes D.L. et al. 2003. A noninva-sive technique for determining sex of live adult North American sturgeons//Environ. Biol. Fish. V. 68. №4. P. 333-338.
Williot P. 2002. Reproduction // Esturgeons et caviar. Paris: Tec et Doc. (Aquaculture-Pisciculture). P. 63-90.
Краснодарский научно-исследовательский институт рыбного хозяйства - КрасНИИРХ, Краснодар
*E-mail: fnd62@mail.ru
Поступила в редакцию 31.12.2004 г.(УДК УДК 597.442.591.4)
Пожалуйста, уважайте труд этих людей и помните о законе РФ «Об авторском праве и смежных правах»
Добавить комментарий